



|
|
|
|
ÉCOSYSTÈMES MARINS: Résultats 2011 - 2014
- Quelle est la dynamique des éléments nutritifs et du carbone organique dissous et particulaire (transport
latéral et vertical y compris les apports continentaux, processus biologiques et chimiques de transformation), et comment sont-ils affectés par les changements en cours (Tremblay, Babin)?
P lusieurs de nos travaux ont porté sur l'importance des apports de nutriments fluviaux dans l'océan Arctique, où la couche de surface se trouve généralement appauvrie en azote (un élément essentiel pour les producteurs primaires). Comme l'océan Arctique reçoit d'énormes apports en eau douce des grands fleuves qui le bordent (10% des apports de l'océan mondial alors que son volume n'en fait que 1%) et le débit de ces fleuves est en augmentation, nous suspections qu'ils pourraient être une source additionnelle de nutriments pour le phytoplancton marin. Or, après avoir assemblé une base de données contenant les mesures historiques de nutriments dans les principaux fleuves et voies d'entrée océaniques, nous avons estimé que le nitrate d'origine fluviale contribuait pour moins de 1% à la production primaire nouvelle annuelle de l'océan Arctique dans son ensemble (Le Fouest et al. 2013a). A l'été par contre, il appert que les apports d'azote organique dissous et la photo-ammonification sur les plateaux peu profonds sont des sources importantes d'azote à proximité des fleuves (Xie et al. 2012). Ces sources pourraient prendre de l'importance avec la fonte du pergélisol dans les bassins versants. Un exercice de modélisation 1-D indique que la prise en compte de la photo-ammonification de composés organiques labiles est nécessaire pour expliquer les niveaux de production primaire observés dans les premiers mètres de la colonne d'eau sur le plateau adjacent au fleuve Mackenzie (Le Fouest et al. 2013b).
Une étude détaillée de cette région durant Malina a révélé que la contribution du nitrate fluvial à la production nouvelle sur l'ensemble du plateau ne dépasse pas 5% même lors d'une année à fort débit. Cette contribution est beaucoup plus importante à l'échelle plus fine de l'estuaire, où l'activité bactérienne recycle l'azote consommé par le phytoplancton et supporte une forte production régénérée (Tremblay et al. sous presse). Nous avons également mis en évidence que la rencontre des eaux fluviale déficitaires en phosphore avec les eaux océaniques déficitaires en azote était nécessaire au maintien d'une productivité relativement forte en zone estuarienne. Ces déficits respectifs résultent de l'ensemble des processus biogéochimiques qui conditionnent les eaux du fleuve le long du bassin versant et les eaux océaniques qui entrent en Arctique par la mer de Béring. En altérant ces processus et modifiant les ratios azote : phosphore des différentes eaux sources, les changements climatiques pourraient affecter la distribution spatiale et l'intensité de la productivité biologique dans les eaux côtières et du large (Tremblay et al., Progress in Oceanography, soumis).
Les fleuves arctiques sont, aussi, une source très importante de carbone organique dissous et particulaire qui pourrait aller en s'amplifiant avec le dégel du pergélisol. L'ampleur (contribution au budget de C de l'océan Arctique (OA) et la dynamique (variations saisonnières et multi-annuelles) de ce flux, et le devenir de ce carbone dans l'océan Arctique (transport, dégradation, enfouissement, entrée dans la chaine trophique) sont très mal connus. Grâce aux nombreuses et détaillées données d'optique du projet Malina (e.g. Matsuoka et al. 2012, Antoine et al. 2013, Bélanger et al. 2013a), nous avons pu développer des algorithmes qui permettent de suivre l'export de COP et de COD fluviatiles vers l'océan Arctique et son advection en surface à partir de données satellitales de radiométrie visible et proche-infrarouge (« couleur de l'océan » ; Doxaran et al. 2012, Fichot et al. 2013, Matsuoka et al. 2013). Les relations entre propriétés optiques qui sous-tendent ces algorithmes ont été en outre et autant que possible validées à l'échelle pan-Arctique (Matsuoka 2014). Ces algorithmes sont maintenant utilisés pour établir des séries temporelles pan-arctiques de l'export annuel de COP et de COD depuis 1997 (Matsuoka et al. en prép., Doxaran et al. en prép.). Les premiers résultats laissent entrevoir une augmentation significative de l'export de COP et une stabilité de l'export de COD par les fleuves arctiques.
Environ 23 Mt de carbone organique sont transférés du continent vers l'AO par année par voie fluviale ou aérienne, ou par érosion côtière (11 Mt de COD selon Amon 2004, et 12 Mt de COP selon Stein & Macdonald 2004).20 à 50% du COD sont exportés hors de l'AO vers l'Atlantique nord (Amon 2004) alors qu'une faible fraction est photo-oxydée pour produire du CO et du CO2 (Song et al. 2013) Le reste est essentiellement minéralisé par l'activité bactérienne. Contrairement à ce qui était admis jusqu'à maintenant, il semble que le COP d'origine terrigène n'est pas plus réfractaire à la dégradation que le COP produit par le phytoplancton dans l'océan (Rontani et al. 2012). Aussi, bien que l'appauvrissement chronique en nutriments inorganiques dans plusieurs régions de l'OA pourrait limiter l'activité bactérienne, c'est bien le COD (et le COP) qui contrôle ce processus (Ortega-Retuerta et al. 2012). Malgré la faible contribution relative des sources terrigènes au bilan de carbone organique de l'OA (la production nette communautaire est de 345 Mt par an selon Codispoti et al. 2004), près de l'embouchure des grands fleuves arctiques, la production biologique est majoritairement hétérotrophe et largement fondée sur l'assimilation du carbone organique d'origine terrigène (Ortega-Retuerta et al. 2012).
- Comment la diversité et la structure fonctionnelle des communautés microbiennes sont-elles affectées par les changements en cours dans les biotopes marins (Lovejoy, Massé)?
La biodiversité et la biogéographie des communautés microbiennes dans l'océan mondial sont peu comprises. Nos recherches ont commencé à faire la lumière sur ce sujet dans l'océan Arctique. Nous avons publié plusieurs articles sur la distribution et la phylogénie des taxons suivants : Cercozoa, diatomées, dinoflagellés et petits flagellés (Thaler et Lovejoy 2012, Luddington et al 2012, Potvin et al 2012, Terrado et al 2012, Thaler et Lovejoy 2014), ainsi que sur des groupes particuliers de bactéries (Galand et al 2013, Alonso-Saez et al. soumis) et d'archées (Pedneault et al 2014). Nous avons également examiné les changements dans le temps au sein de l'ensemble de la communauté microbienne des maxima de biomasse de sub-surface dans le golfe d'Amundsen (Comeau et al 2012) et examiné les protistes de la glace de mer (Comeau et al 2013). Nous avons de plus identifié des communautés distinctes de flagellés hétérotrophes dans les trois masses d'eau supérieures de l'Arctique de l'Ouest (Monier et al 2013). En outre, Lovejoy a contribué à des articles sur les distributions bipolaires de bactéries (Ghiglione et al 2013) et les archées (Alonso-Saez et al 2013) et écrit des articles de revue sur les protistes (Lovejoy 2013) et sur les bactéries de l'Arctique (Pedros-Alio et Lovejoy, à l'examen). Les tendances qui se dégagent de la biodiversité et le potentiel de changement dans l'Arctique se reflètent dans la participation de Lovejoy dans l'évaluation de la biodiversité de l'Arctique (ABA) http://www.caff.is/assessmentseries/view_document/233-arctic-biodiversity-assessment-2013 et les comités nationaux et internationaux sur le plan de la surveillance maritime circumpolaire (CBMP).
Parce que la composition des communautés microbiennes répond fortement aux changements des conditions environnementales, on peut aussi interpréter les changements passés de composition de communauté pour en tirer des informations sur les conditions environnementales qui régnaient alors. Cette approche paléocéanographique passe par l'interprétation de biomarqueurs tels que l'IP25 produit par des diatomées qui vivent exclusivement en association avec la glace de mer, et les dinokystes ou autres reliques de phytoplancton, et de proxy géochimiques variés (composition isotopique du carbone, acides gras spécifiques, …). Nous avons mené des études à partir d'archives sédimentaires collectées dans plusieurs régions clés de l'Arctique. Nos résultats ont été publiés dans des revues à fort impact. En particulier, l'analyse de sédiments récoltés en mer du Labrador a montré que la transition abrupte d'un climat froid et rigoureux au cours du Younger Dryas vers des conditions plus clémentes au début de l'Holocène était liée à une augmentation de l'intensité du Gulf Stream par rapport au courant du Labrador (Pierce et al. 2013, Nature Communications). Ce transport accru de masses d'eau chaudes est à l'origine du bouleversement des conditions atmosphériques observées précédemment dans les hautes et basses latitudes. Cette étude a donc permis de démontrer que, contrairement à ce qui était communément admis, ce sont des variations des conditions océaniques de surface qui ont présidé au changement de conditions atmosphérique et non l'inverse. Nos travaux ont aussi montré que les conditions de surface dans la zone ont oscillées entre périodes chaudes faiblement englacées et périodes plus froides ou la glace, transportée
par le courant froid du labrador atteignait le sud de terre neuve (Weckström et al. 2013, QSR ; Sicre et al. 2013, EPSL). En particulier, Sicre et al. ont montré que pendant les périodes froides, le fort englacement de la zone retardait le bloom printanier jusque pendant l'été quand les eaux de surface sont plus chaudes.
- Quelle est la réponse de la production primaire aux changements des propriétés chimiques (nutriments), physiques (température, hydrodynamiques, optiques) de l'océan, de biodiversité et d'englacement (Babin, Tremblay)?
Grâce à un modèle diagnostique de production primaire (PP) marine optimisé à partir des données du projet Malina, nous avons pu évaluer la réponse de la PP aux changements environnementaux actuels en Arctique en utilisant les données de télédétection radiométrique visible-PIR (« couleur de l'océan ») disponibles depuis 1998 (Bélanger et al. 2013b). À l'échelle pan-arctique et sur la période 1998-2010, nous avons estimé une PP annuelle moyenne de 203 Tg C, une valeur deux fois plus faible que celles publiées récemment par d'autres auteurs. Cette différence s'explique par la prise en compte dans notre modèle des propriétés optiques de l'eau de mer très particulières de l'OA. Par ailleurs, la PP annuelle a augmenté d'environ 14% par décennie sur cette même période essentiellement à cause de la réduction de l'étendue de la banquise en été qui permet une pénétration accrue du rayonnement solaire dans l'océan, mais aussi à cause d'une augmentation de la biomasse.
En revanche, cette augmentation est freinée par la réduction du rayonnement solaire incident (sous les
nuages et au-dessus de la surface de l'océan) de 8% par décennie qui résulte d'une augmentation de la nébulosité.Est-ce que les émissions de DMS par le phytoplancton contribueraient à cette augmentation de la nébulosité ?
Dans une étude récentes (Ardyna et al., soumis à GRL), nous avons pu mettre en évidence ce qui semble être un profond bouleversement du fonctionnement de l'écosystème marin arctique, perceptible sur la
période 2008-2012 par télédétection. La phénologie du phytoplancton montre en effet qu'un bloom d'automne causé par le brassage lié au vent est de plus en plus fréquent aux hautes latitudes (Figure 1).
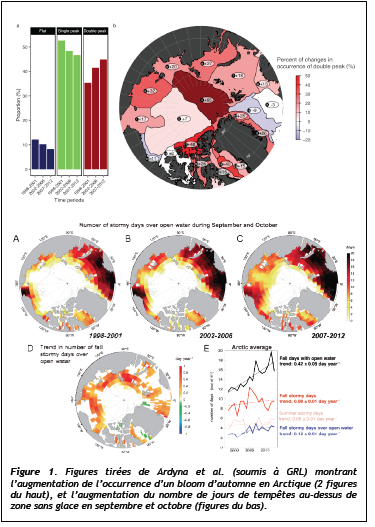
Dans un océan stratifié à l'extrême à cause des apports en eaux douce, le mélange vertical est donc malgré tout possible quand il y a moins de glace de mer. La disparition de la banquise d'été permet non seulement au rayonnement solaire de pénétrer dans l'océan, mais elle accentue le mélange vertical,
source de nouveaux nutriments.
Malgré son fort potentiel, la télédétection de la couleur de l'océan rencontre néanmoins certaines difficultés spécifiques à l'OA : surface englacées néanmoins productives, forte couverture nuageuse à partir du mois d'août, contamination du signal par la glace de mer, et faibles élévations solaires. Par ailleurs, une fraction significative de la PP est réalisée au niveau d'accumulations de biomasse phytoplanctonique autour de 40 à 70 m de profondeur (les « SCM »), une caractéristique de l'OA liée à la forte stratification de ce système.
Ces SCMs ne sont pas « vus » par les satellites car trop profonds. Nous avons pu établir en analysant une vaste base de données in situ panarctique, que la présence de SCM ne faussait que très faiblement nos estimations pan-arctiques et annuelles de PP par télédétection (Ardyna et al. 1013). Par contre, à l'échelle régionale et en août et septembre, l'omniprésence des SCM pourrait conduire à de fortes sous-estimations de la PP. Un tandem d'articles (Martin, Tremblay et al. 2012, 2013 - 44 citations) a permis pour la première fois d'élucider le fonctionnement des SCMs arctiques, montrant qu'ils contribuent fortement à la PP et aux flux azotés en raison de la forte efficacité photosynthétique des algues en basse lumière.
En Arctique, comme ailleurs dans l'océan, on ne peut aborder le rôle de la lumière et des nutriments dans les variations de la production primaire sans considérer les couplages avec les processus hydrodynamiques. Aussi, l'Arctique présente de fortes spécificités régionales liées à la physionomie du bassin et aux conditions aux limites (plateau continental couvrant la moitié du bassin, nombreux passages à travers des archipels complexes, connexions avec le Pacifique et l'Atlantique, forts apports en eau douce, dynamique complexe de la banquise). Nous avons récemment examiné de plus près le régime de PP dans certaines régions comme la Mer de Beaufort et la Baie de Baffin. Contrairement aux attentes "intuitives", nous avons montré que le déclin des glaces mène à une diminution de la PP dans une région où l'eau de fonte s'écoule hors de l'Arctique (nord de la Baie de Baffin), mais à une augmentation de la PP dans une région intérieure (Mer de Beaufort ; Bergeron et Tremblay 2014). Cette augmentation est causée soit par l'approfondissement des couches profondes de phytoplancton, qui bénéficient d'un éclairement accru, soit par l'upwelling, qui peut conduire à des pics de productivité spectaculaires en marge des plateaux peu profonds (Tremblay et al. 2011, Geophysicial Research Letters + choix de l'éditeur dans Science – 36 citations). Sur ces mêmes plateaux, nous avons démontré pour la première
fois l'existence de taux modérés de fixation de l'azote gazeux, processus qu'on croyait jusqu'alors incompatible avec les températures froides. Cette fixation redresse partiellement le bilan azoté de l'océan Arctique (Blais et al. 2012, Global Biogeochemical Cycles 26, repris dans Nature). Plusieurs de ces résultats ont été publiés dans des revues à fort impact et ont fait l'objet d'une couverture médiatique par des journaux influents tels que The Economist et The Financial Times.
Pour approfondir encore plus notre compréhension des variations de PP en Arctique, nous avons examiné les propriétés photosynthétiques du phytoplancton lors du projet Malina. Nos résultats révèlent une acclimatation extrême aux très faibles éclairements qui règnent en Arctique (Huot et al. 2012). Ces caractéristiques du phytoplancton arctique lui permettent de croitre rapidement malgré des conditions défavorables de lumière et de température. Elles font maintenant l'objet d'expériences en laboratoire pour mieux comprendre les mécanismes qui sous-tendent cette photoacclimatation extrême, et comment ils interviennent dans la succession des diatomées au printemps.
- Comment les modifications de régime de production primaire affectent-elles les flux de carbone, et le transfert d'énergie et de matière au niveau des échelons suivants de la chaine trophique (Fortier, Maps, Babin)?
Nous avons réalisé plusieurs travaux sur les flux de carbone organique particulaire arctiques (Mer de Beaufort, de Laptev et de Sibérie orientale, Archipel canadien, Baie d'Hudson, Détroit de Fram). Des séries temporelles de longue durée permettent de suivre l'évolution de la capacité de l'Océan Arctique à séquestrer le CO2 et, surtout, d'évaluer l'impact des variations da la production primaire sur le couplage pélagos-benthos et les réseaux trophiques marins arctiques. Les conclusions d'une douzaine de publications primaires ont contribué à ce que l'on peut considérer comme la synthèse la plus à jour des flux de carbone marins arctiques. (Figure 2)
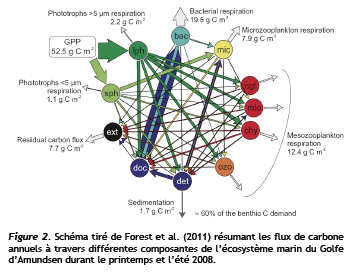
Une étude similaire pour le plateau du Mackenzie a récemment été publiée (Forest et al. 2013). Par ailleurs, nous avons détecté les premiers symptômes potentiels de l'altération prévue de la composition des maillons supérieurs du réseau pélagique. Ainsi, en Mer de Beaufort, la croissance des effectifs du grand copépode herbivore Calanus glacialis de 2007 à 2011 a coïncidé avec l'intensification de l'upwelling dans le Golfe d'Amundsen (Tremblay et al. 2011, Geophysical Research Letter, doi:10.1029/2011GL048825; Suzuki et al. en préparation). De même, des conditions plus favorables de production primaire et de température pourraient sous-tendre l'invasion récente de la Mer de Beaufort par le lançon du Pacifique, un compétiteur potentiel de la morue arctique (Falardeau-Côté et al. 2014, ICES Journal Marine Sciences. DOI: 10.1093/icesjms/fst221). Enfin, les impacts potentiels d'une modification des régimes de production des algues de glace et du phytoplancton sur les migrations verticales et la reproduction du zooplancton ont été explorés sur un cycle annuel complet en
Mer de Beaufort (Darnis and Fortier 2014 Journal of Plankton Research doi:10.1093/plankt/fbu035).
L'ensemble de ces résultats contribue directement aux Études Intégrées Régionales des Impacts du RCE Arctic-Net.
Les méthodes numériques (analyse de bases de données et modèles informatiques) permettent d'aborder efficacement l'écologie du zooplancton des mers sub-Arctiques et Arctiques. Le zooplancton est une interface essentielle entre les producteurs primaires unicellulaires et les niveaux supérieurs du réseau trophique, entre la variabilité de l'environnement physique et les propriétés des écosystèmes. Un modèle de la biologie de l'espèce clé de copépode du golfe du Saint-Laurent, Calanus finmarchicus, a été couplé à un modèle de circulation régionale (Maps et al. 2011, doi : 10.1016/j.jmarsys.2011.04.004). Il constitue un point de départ pour nos études par modélisation du zooplancton arctique. Ce travail a révélé le « circuit Calanus » constitué de structures hydrodynamiques distinctes reliées entre elles par la phénologie et le comportement de C. finmarchicus et sur lequel repose sa pérennité dans ce système très dynamique. L'importance des copépodes dans les écosystèmes marins Arctiques repose en particulier sur leur faculté de s'affranchir des conditions hivernales défavorables pendant une longue période de diapause. Ils peuvent ainsi rendre disponible sur une longue période à leurs prédateurs la biomasse synthétisée en été par le phytoplancton. Un travail de méta-analyse de données publiées à permis de démontrer que le métabolisme des lipides pouvait fournir un mécanisme de contrôle de la diapause chez les espèces arctiques mais aussi à travers une quinzaine d'espèces (Maps et al. 2013,
doi : 10.1093/plankt/fbt100). Ce travail a bénéficié du développement en parallèle d'un cadre numérique basé sur les principes premiers de la thermodynamique et de la biologie du zooplancton qui permet de répondre à des problématiques globales telles que les patrons de biodiversité des copépodes (Record et al. 2012, doi :10.1007/s00442-012-2313-0) ou les mécanismes de co-existance au sein des communautés de plancton (Record et al 2013a, doi : 10.1093/icesjms/fst049). Le niveau d'organisation des communautés est idéal pour réconcilier les approches biogéochimiques et par espèces cibles qui ont cours en parallèle depuis une vingtaine d'années (Record et al. 2013b, doi : 10.1016/j.ecolmodel.2013.03.018). Il s'agit aussi d'une étape inévitable vers une meilleure représentation des niveaux trophiques intermédiaires dans les modèles « end to end » (de l'environnement aux espèces apicales) sensés développer nos capacités prévisionnelles en écologie marine. Les recherches en cours dans le laboratoire consistent en un mélange d'expériences de laboratoires (Maîtrise en cours sur la prédation intraguilde chez les copépodes arctiques) et d'observations de terrain fournissant des
paramètres et des données de validation à des expériences de modélisation numériques d'écologie théoriques et appliquées (e.g. Maps et al. 2013, doi : 10.1093/icesjms/fst021).
- Est-ce que les changements de production primaire rétroagissent sur le climat via la production de composés sulfurés (Levasseur, Massé, Babin)?
Une première revue de la dynamique du diméthylsulfure (DMS) en Arctique a été publiée dans le journal Nature Geoscience (Levasseur 2014). Cette revue a mis en évidence la diversité des sources potentielles de DMS pour l'atmosphère en Arctique et a permis d'explorer comment le réchauffement en cours pourrait altérer ces sources. Nos travaux dans l'Arctique canadien ont d'ailleurs porté sur deux de ces sources potentielles et encore non-quantifiées : les floraisons sous la glace et les mares de fonte qui peuvent couvrir jusqu'à 80% de la surface de la banquise pendant la période de fonte. Le suivie de deux floraisons sous la glace dans le Détroit de Barrow, Arctique canadien, lors des printemps 2010 et 2011 a révélé des concentrations élevées en DMS (jusqu'à 11 nmol L1) et en diméthylsulfoniopropionate (DMSP ; jusqu'à 185 nmol L-1), le précurseur algal du DMS (Galindo et al. 2014a,b). Ces valeurs se comparent avantageusement à celles mesurées lors des floraisons prenant place à la marge de la glace en Arctique, mais la présence du couvert de glace pourrait réduire considérablement la ventilation du DMS produit vers l'atmosphère. Les mares de fonte ont également fait l'objet d'un suivi temporel au cours de l'été 2012 révélant des concentrations atteignant 11 nM. Il s'agit d'une observation des plus importantes considérant que cette source de DMS est directement en contact avec l'atmosphère. Nous tentons présentement de déterminer les mécanismes responsables de la formation de ce réservoir de DMS et de déterminer la fraction de ce réservoir qui est éventuellement ventilée vers l'atmosphère. On observe généralement une faible correspondance entre les concentrations de DMS dans les océans et la biomasse phytoplanctonique. Pour cette raison, les estimations de la biomasse microalgale à partir des capteurs satellitaires ne peuvent pas être directement converties en DMS. Nous explorons, depuis l'automne 2013, les différentes possibilités d'effectuer cette conversion, mais de façon indirecte. Deux voies semblent particulièrement prometteuses, soit l'utilisation de la relation généralement observée entre les concentrations de DMS et la dose de radiation solaire journalière aux faibles et moyennes latitudes, de même que l'utilisation d'algorithmes permettant de discriminer les principaux groupes phytoplanctoniques (forts versus faibles producteurs de DMSP). Ces travaux permettront de diagnostiquer les concentrations et émissions de DMS à une meilleure résolution temporelle et spatiale que celle obtenue à partir des climatologies et ce tant à l'échelle globale que régionale (ex. Arctique).